Para su tesis doctoral,
Kristoffer Krogerus, ha estado investigando las propiedades relacionadas con el sabor y el estrés de los híbridos de levadura de cerveza. Se sabe desde hace algún tiempo que la
levadura lager (
Saccharomyces pastorianus) es en realidad una especie híbrida, y que uno de los padres fue la bien conocida
levadura ale Saccharomyces cerevisiae . En 2011, se descubrió el otro lado de la familia,
Saccharomyces eubayanus . Este descubrimiento ha permitido la caracterización mejorada de las levaduras lager, y también ha abierto la posibilidad de crear nuevas cepas de levadura lager a medida. Esto es posible mediante el apareamiento de cepas seleccionadas de las dos especies parentales.
Esto es exactamente lo que ha estado haciendo durante el año pasado, y se complace en anunciar que ha publicamos sus primeros resultados (Nuevas cepas de levadura lager generadas por hibridación interespecífica) en el
Journal of Industrial Microbiology and Biotechnology. Se acopló una cepa de cerveza de producción fuertemente floculante (de una fábrica de cerveza en el Reino Unido) con S. eubayanus, para producir híbridos de levadura lager que se desempeñaron mejor que las cepas parentales y heredaron propiedades beneficiosas de ambas. Esto abrirá la posibilidad de producir una variedad de nuevas cepas de levadura lager, con, por ejemplo, una producción de sabor interesante y una mayor tolerancia al estrés. Ya tenemos muchas nuevas combinaciones híbridas interesantes que esta deseando caracterizar. Publicará más detalles en una publicación posterior, pero mientras tanto, siéntase libre de leer la publicación si está interesado, ¡es
Open Access!
Resumen:
El híbrido interespecífico Saccharomyces pastorianus es la levadura más utilizada en fermentaciones de cervecería en todo el mundo. Aquí generamos híbridos de levadura de novo lager mediante el apareamiento de una cepa de Saccharomyces cerevisiae domesticada y fuertemente floculenta con la Saccharomyces eubayanus.tipo de cepa Los híbridos se caracterizaron con respecto a las cepas parentales en una fermentación de mosto realizada a temperaturas típicas para la elaboración de cerveza lager (12 ° C). Las cervezas resultantes se analizaron para determinar los compuestos de azúcar y aroma, mientras que las levaduras se analizaron para determinar su capacidad de floculación y capacidad de transporte de α-glucósido. Estos híbridos heredaron propiedades beneficiosas de ambas cepas parentales (criotolerancia, utilización de maltotriosa y fuerte floculación) y mostraron un vigor híbrido aparente, fermentando más rápido y produciendo cerveza con mayor contenido de alcohol (5,6 vs 4,5% ABV) que los padres. Los resultados sugieren que la hibridación interespecífica es adecuada para la producción de nuevas cepas de levadura lager no GM con propiedades únicas y ayudará a dilucidar la historia evolutiva de la levadura lager industrial.
Nuevas cepas de levadura lager generadas por hibridación interespecífica.
Autores y afiliaciones: Kristoffer Kroger, Frederico Magalhães, Virve Vidgren, Brian Gibson
15 de febrero de 2015
Resumen:
El híbrido interespecífico Saccharomyces pastorianus es la levadura más utilizada en fermentaciones de cervecería en todo el mundo. Aquí generamos híbridos de levadura de novo lager mediante el apareamiento de una cepa de Saccharomyces cerevisiae domesticada y fuertemente floculenta con la Saccharomyces eubayanus.tipo de cepa Los híbridos se caracterizaron con respecto a las cepas parentales en una fermentación de mosto realizada a temperaturas típicas para la elaboración de cerveza lager (12 ° C). Las cervezas resultantes se analizaron para determinar los compuestos de azúcar y aroma, mientras que las levaduras se analizaron para determinar su capacidad de floculación y capacidad de transporte de α-glucósido. Estos híbridos heredaron propiedades beneficiosas de ambas cepas parentales (criotolerancia, utilización de maltotriosa y fuerte floculación) y mostraron un vigor híbrido aparente, fermentando más rápido y produciendo cerveza con mayor contenido de alcohol (5,6 vs 4,5% ABV) que los padres. Los resultados sugieren que la hibridación interespecífica es adecuada para la producción de nuevas cepas de levadura lager no GM con propiedades únicas y ayudará a dilucidar la historia evolutiva de la levadura lager industrial.
Introducción
Pale lager es el estilo de cerveza más popular en todo el mundo. Todas las cervezas de este tipo se producen mediante la fermentación del mosto de malta a baja temperatura con la especie de levadura híbrida interespecífica Saccharomyces pastorianus [14]. La formación de híbridos interespecíficos es común entre las especies estrechamente relacionadas del género Saccharomyces, y la levadura lager S. pastorianus es un híbrido natural entre S. cerevisiae y el recién descubierto S. eubayanus [21]. Las especies híbridas son comunes en ambientes antropogénicos y no se limitan a la levadura, ya que las especies híbridas a menudo exhiben cualidades superiores a ambas cepas parentales [22, 33]. Las cervezas Lager han ganado popularidad debido a su perfil de sabor "limpio", es decir, a la falta de aroma afrutado / floral derivado del éster, que resulta principalmente de la fermentación a bajas temperaturas. Las cepas de Ale (S. cerevisiae) se han seleccionado con el tiempo por sus propiedades de elaboración beneficiosas, como la floculación, la utilización del azúcar y el perfil de sabor, pero requieren temperaturas más cálidas (> 15 ° C) para una fermentación óptima [40]. En contraste, S. eubayanus prospera a las bajas temperaturas responsables del perfil de aroma típico de la cerveza lager, pero carece de la capacidad de utilizar maltotriose (el azúcar más abundante en el mosto después de la maltosa) y muestra una pobre floculación [13].
No se conocen los orígenes de la levadura lager, pero es posible que el evento de hibridación original se deba a que S. eubayanus contaminó una fermentación tradicional de cerveza con S. cerevisiae [14]. El grupo S. pastorianus consta de al menos dos linajes distintos (Saaz y Frohberg), que pueden haber surgido de forma independiente [11, 23] o mediante un evento de hibridación común [42]. Si bien estos dos grupos comparten muchas características comunes, difieren funcionalmente en varios aspectos, como la utilización de la maltotriosa y la tolerancia al frío [13, 34]. Estas diferencias funcionales parecen reflejarse en la diferencia genómica entre los grupos (las cepas de Saaz han retenido proporcionalmente más ADN derivado del progenitor de S. eubayanus, mientras que las cepas de Frohberg han retenido proporcionalmente más ADN del progenitor de S. cerevisiae). Debido a que las levaduras lager están restringidas solo a estos dos grupos genéticamente distintos, la diversidad genética entre ellos es pobre. Sin embargo, a través de la selección de cepas individuales de S. eubayanus y S. cerevisiae con características específicas deseables, y la posterior hibridación de estas cepas, puede ser posible producir nuevas cepas de levadura hechas a la medida y disponibles de inmediato para uso industrial.
El uso de la hibridación interespecífica para generar cepas de cerveza y levadura de vino con producción mejorada de aroma se ha descrito [1, 12, 38, 39]. Aquí describimos, por primera vez, el uso de apareamiento en masa para generar híbridos de levadura lager de novo lager a partir de una cepa de ale de S. cerevisiae fuertemente floculante domesticada y la cepa de tipo S. eubayanus. Los híbridos se caracterizaron con respecto a las cepas parentales en una fermentación de mosto realizada a temperaturas típicas para la levadura lager, y la cerveza resultante se analizó en cuanto al contenido de azúcar y la levadura se analizó para determinar su capacidad de floculación. El objetivo de este estudio fue demostrar la generación de nuevas cepas de levadura lager con propiedades fisiológicas heredadas de ambas cepas parentales. Esta técnica tiene el potencial de aumentar considerablemente la diversidad de cepas de levadura disponibles para la elaboración de cerveza lager [14]. Los híbridos de levadura generados con esta técnica también pueden verse como no GM, y su uso no está impedido por la legislación o la opinión pública [6].
Materiales y métodos
Cepas de levadura
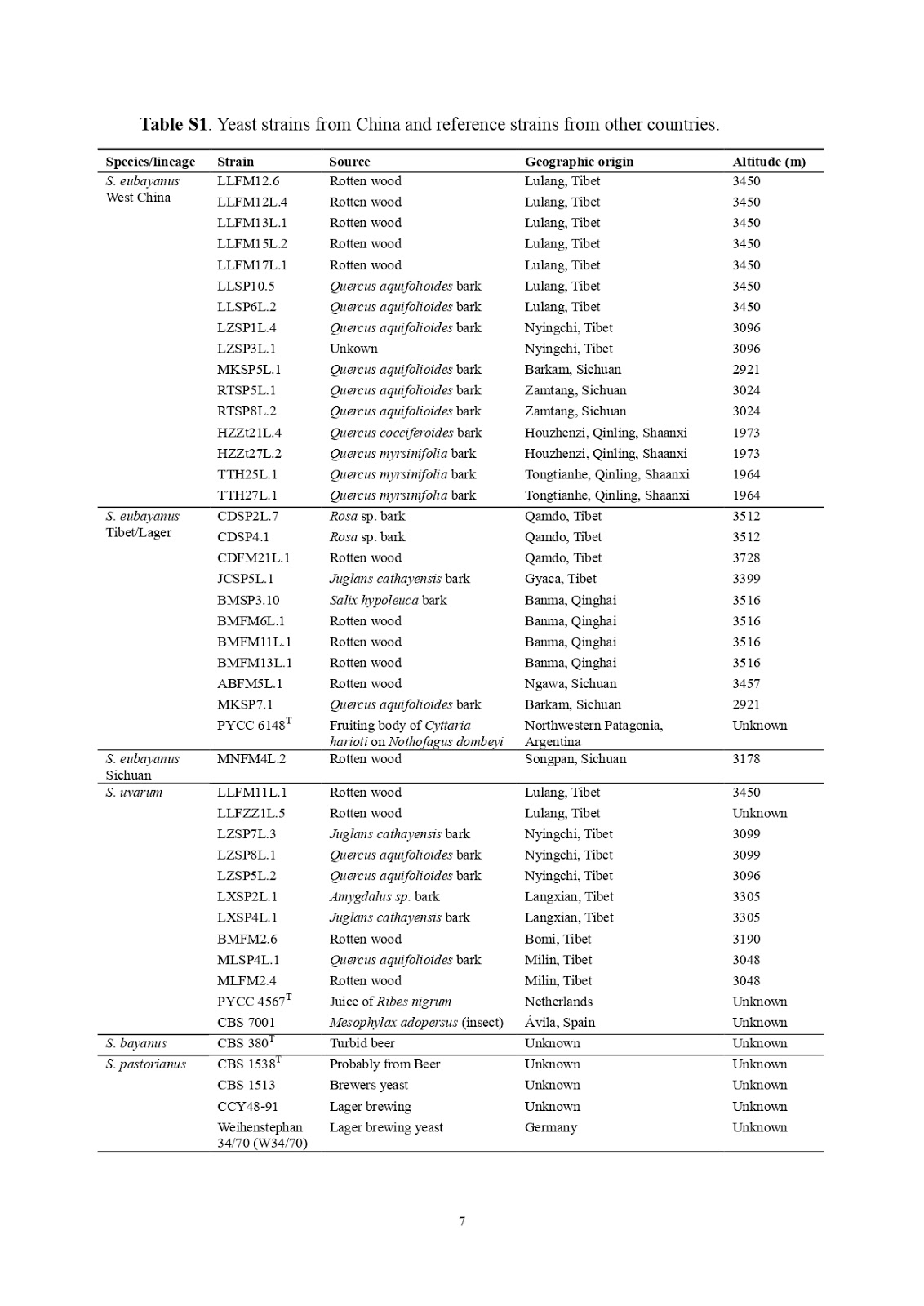
Las dos cepas parentales fueron S. cerevisiae VTT-A81062 (VTT Culture Collection, Finlandia), una cepa de levadura de cerveza originaria de una cerveza de cerveza proveniente del Reino Unido, y la cepa de tipo S. eubayanus VTT-C12902 (VTT Culture Collection, Finlandia ; depositado como CBS12357 en CBS-KNAW Fungal Biodiversity Center). Las cuatro cepas híbridas (A81062 × C12902) que se eligieron para una caracterización adicional se denominaron H1-H4. Se seleccionaron mutantes auxótrofos naturales (lys y ura) de las cepas parentales en placas de agar con ácido α-aminoadípico y ácido 5-fluoroorótico, respectivamente [4, 50]. La auxotrofia se confirmó por la incapacidad de crecer en un medio de agar de selección mínima (0,67% de Base de Nitrógeno de Levadura sin aminoácidos, 3% de glicerol, 3% de etanol y 2% de agar).
Esporulación
Para la generación de ascosporas, los mutantes auxotróficos de las cepas parentales se cultivaron primero durante la noche en medio YPM (1% de extracto de levadura, 2% de peptona, 4% de maltosa) a 20 ° C. Luego se inoculó la levadura en medio de pre-esporulación (0,8% de extracto de levadura, 0,3% de peptona, 10% de glucosa) a una OD600 de 0,3 y se dejó crecer durante 20 horas a 20ºC. Luego se lavó la levadura con acetato de potasio al 1% y se colocó una suspensión espesa sobre agar de esporulación (acetato de potasio al 1%, lisina y uracilo 10% / l, agar de 2%). La levadura se dejó esporular durante 7 días a 25ºC. La eficacia de la esporulación se calculó contando la frecuencia de ascosporas teñidas con verde de malaquita [27]. La viabilidad de las esporas se calculó mediante la disección de ascosporas tratadas con Zymolyase 100T (US Biological, EE. UU.) En agar YPD con un micromanipulador [43].
Generación de híbridos interespecíficos.
Los híbridos interespecíficos entre un ura-aislado de S. cerevisiae A81062 y un lis-aislado de S. eubayanus C12902 se produjeron primero generando ascosporas de los mutantes auxotróficos como anteriormente. Las ascosporas se rasparon del agar en 1 ml de H2O purificada por ósmosis inversa estéril en tubos Eppendorf de 2 ml. Los tubos se centrifugaron a 5.000 xg durante 5 minutos y se eliminó el sobrenadante. Las paredes de Ascus se digirieron mediante la adición de 50 μl de 1 mg / ml de Zymolyase 100T y la incubación a 30 ° C durante 20 min. Luego se agregaron 200 μl de H2O estéril y las células se resuspendieron mediante agitación con vórtex. Se transfirieron 100 μl de las suspensiones resultantes de ambas cepas parentales, con marcadores auxotróficos complementarios, a 1 ml de medio YPM en un tubo Eppendorf estéril de 2 ml. Los tubos se agitaron e incubaron estáticamente a 25ºC durante 7 días. Después de la incubación, los tubos se centrifugaron a 5.000 xg durante 5 minutos y se eliminó el sobrenadante. Se agregaron 500 μl de medio de inanición (0,1% de extracto de levadura y 0,1% de glucosa) y los tubos se incubaron durante al menos 2 horas a temperatura ambiente. Los tubos se agitaron en vórtex, después de lo cual se midió la concentración celular aproximada de la suspensión resultante con un NucleoCounter YC-100 (ChemoMetec, Dinamarca) y se distribuyeron alícuotas de 100 μl en agar de selección mínima (sin uracilo o lisina). Las placas se incubaron a 25 ° C y aparecieron colonias prototróficas (es decir, híbridos potenciales) después de 3 a 7 días. Las colonias se contaron y se purificaron por repilación en agar de selección mínima.
Confirmación del estado híbrido por PCR y RFLP
El estado híbrido de los aislamientos se confirmó mediante la amplificación de rDNA-PCR (ITS1, 5.8S e ITS2) utilizando los cebadores ITS1 (5′-TCCGTAGGTGAACCTGCGG-3 ') y ITS4 (5′-TCCTCCGCTTATTGATATGC-3 ′) y ITS4 (5′-TCCTCCGCTTATTGATATGC-3') la enzima de restricción HaeIII (New England BioLabs, EE. UU.) como se describió anteriormente [31]. La identificación se basó en el número de fragmentos de restricción generados por la digestión enzimática. S. eubayanus produjo un patrón de 3 bandas (490, 225, 140 pb), S. cerevisiae un patrón de 4 bandas (320, 225, 180, 140 pb), mientras que los híbridos exitosos produjeron un patrón con las 5 bandas (490, 320, 225, 180 y 140 pb).
La amplificación del gen FSY1 específico de S. eubayanus (tamaño del amplicón 228 pb) y el gen MEX67 específico de S. cerevisiae (tamaño del amplicón 150 pb) también se realizó en el ADN extraído de las cepas híbridas utilizando los cebadores SeubF3 (5'- GTCCCTGTACCAATTTAATATTGCGC-3 ′), SeubR2 (5′-TTTCACATCTCTTAGTCTTTTCCAGACG-3 ′), ScerF2 (5'GP.) [28] y Pengelly y Wheals [30]. Los híbridos fueron identificados por la presencia de ambos genes.
Confirmación del estado híbrido por PFGE
Las cepas de levadura se propagaron en YPM a 20 ° C a un OD600> 1 y luego se recolectaron por centrifugación (3,000 × g, 5 min, 4 ° C). Los sobrenadantes se decantaron y las células se resuspendieron en 10 ml de EDTA 50 mM a 4ºC (pH 8). Las concentraciones celulares se determinaron con un Nucleocounter® YC-100 ™ (ChemoMetec) y se colocaron 1,2 x 108 células en cada tapón de muestra. Los tapones de muestra se prepararon con el kit de conectores de ADN genómico CHEF para levadura (Bio-Rad) de acuerdo con las instrucciones del fabricante.
Los tapones de muestra se cargaron en los pocillos de un gel de agarosa (Bio-Rad) certificado en el campo del pulso al 1,0%. El PFGE se realizó a 14 ° C en 0,5 x tampón TBE [89 mMTris, ácido bórico 89 mM, EDTA 2 mM (pH 8)]. Se utilizó un sistema de electroforesis de campo pulsado CHEF Mapper XA (Bio-Rad) con los siguientes ajustes: 6 V / cm en un ángulo de 120 °, la longitud del pulso aumenta linealmente de 26 a 228 s, y el tiempo total de ejecución de 38 h. Se usó una preparación de marcador de cromosoma comercial de la cepa YNN295 de S. cerevisiae (Bio-Rad) para la calibración de la masa molecular. Después de la electroforesis, los geles se tiñeron con bromuro de etidio y se escanearon con el sistema de imágenes Gel Doc XR + (Bio-Rad).
Contenido de ADN por citometría de flujo.
La citometría de flujo se realizó en las cepas de levadura esencialmente como se describe por Haase y Reed [16]. Las células se cultivaron durante la noche en medio YPD (extracto de levadura al 1%, peptona al 2%, glucosa al 2%) y se lavaron aproximadamente 1 x 107 células con 1 ml de tampón citrato 50 mM. Las células se fijaron luego con etanol frío al 70% y se incubaron a temperatura ambiente durante 1 h. Las células se lavaron luego con tampón de citrato 50 mM, se resuspendieron en tampón de citrato 50 mM que contenía 0,25 mg ml-1 ARNasa A y se incubaron durante la noche a 37ºC. Luego se agregó 1 mg mL-1 de proteinasa K, y las células se incubaron durante 1 hora a 50 ° C. Luego, las células se tiñeron con SYTOX Green (2 μM; Life Technologies, EE. UU.) Y su contenido de ADN se determinó utilizando un citómetro FACSAria (Becton-Dickinson). Los contenidos de ADN se calcularon comparando las intensidades de fluorescencia con las de las cepas de referencia haploides de S. cerevisiae (CEN.PK113-1A) y diploides (CEN.PK). Las mediciones se realizaron en cultivos de levadura independientes duplicados y se recolectaron 100,000 eventos por muestra durante la citometría de flujo.
Caracterización de cepas híbridas.
Se seleccionaron cuatro híbridos seleccionados al azar (H1-H4) y las cepas parentales para una caracterización adicional en una fermentación de mosto a pequeña escala realizada a 12 ° C. La levadura se propagó esencialmente como se describe anteriormente [19], con el uso de una fermentación de 'Generación 0' antes de las fermentaciones experimentales reales. Las fermentaciones experimentales se llevaron a cabo por duplicado, en recipientes de fermentación cilindrocónicos de acero inoxidable de 2 l, que contenían 1,5 l de medio de mosto. El mosto se produjo en VTT Pilot Brewery a partir de malta de cebada y malta de trigo, y contenía un contenido de extracto de 12.0 ° Plato (59 g de maltosa, 19 g de maltotriosa, 16 g de glucosa y 4.6 g de fructosa por litro) y nitrógeno amino libre ( FAN) contenido de 269 mg L − 1. La levadura se inoculó a una tasa de 4 g de levadura fresca por litro de mosto (correspondiente a 16 × 106 células viables ml-1). El mosto fue oxigenado a 18 mg L-1 antes de lanzarlo. Las fermentaciones se llevaron a cabo a 12 ° C durante 11 días, o hasta que no se observó ningún cambio en el extracto residual durante 24 h. Las muestras de mosto se extrajeron regularmente de los recipientes de fermentación con una jeringa y se colocaron directamente en hielo, después de lo cual la levadura se separó del mosto de fermentación por centrifugación (9,000 × g, 10 min, 1 ° C).
La floculación de las cepas de levadura se evaluó utilizando un ensayo de Helm modificado esencialmente como lo describen D’Hautcourt y Smart [9]. Los cultivos recuperados de la fermentación se lavaron dos veces con EDTA 0,5 M (pH 7) para romper los agregados celulares y luego se diluyeron a un OD600 de 0.4. La floculación se analizó lavando primero los gránulos de levadura con una solución de CaCl2 4 mM y resuspendiéndolos en 1 ml de solución de floculación que contenía CaCl2 4 mM, 6,8 g / L de acetato de sodio, 4,05 g / L de ácido acético y 4% (v / v) de etanol. (pH 4.5). Las células de levadura en tubos de control se resuspendieron en EDTA 0,5 M (pH 7). Después de un período de sedimentación de 10 min, se tomaron muestras (200 μL) justo debajo del menisco y se dispersaron en EDTA 10 mM (800 μL). Se midió la absorbancia a 600 nm y se determinó el porcentaje de floculación a partir de la diferencia de absorbancia entre los tubos de control y de floculación.
La absorción de maltosa y maltotriosa por las cepas de levadura se ensayó primero cultivándolas en medio YPM a 20ºC. Las levaduras generalmente se recolectaron a una OD600 nm entre 4 y 8 (es decir, a 2 ± 1 mg de levadura seca mL − 1) por centrifugación, se lavaron con agua helada y luego con tartrato de tris 0.1 M helado (pH 4.2) y finalmente se resuspendió en el mismo tampón a una concentración de 200 mg de levadura fresca mL-1. Las tasas de transferencia cero de [U-14C] -maltosa y [U-14C] -maltotriosa absorción se determinaron a 20 ° C con sustrato 5 mM en tartrato de 0,1 M-Tris (pH 4.2) como se describió anteriormente [24], con reacción tiempo de 1 min. [U-14C] -maltose (ARC 488) y [U-14C] -maltotriose (ARC 627) fueron de American Radiolabeled Chemicals Inc. (St. Louis, MO, EUA). La [U-14C] -maltotriose se repurificó antes de su uso como se describe por Dietvorst et al. [10].
Análisis de mosto y cerveza.
La gravedad específica, el nivel de alcohol y el pH de las muestras se determinaron a partir de las muestras de fermentación centrifugadas y desgasificadas utilizando un medidor de densidad Anton Paar DMA 5000 M (Anton Paar GmbH, Austria) con módulos Alcolyzer Beer ME y pH ME (Anton Paar GmbH, Austria) .
El contenido de masa seca de levadura de las muestras (es decir, levadura en suspensión) se determinó lavando los gránulos de levadura obtenidos por centrifugación dos veces con 25 ml de H2O desionizada y luego suspendiendo la levadura lavada en un total de 6 ml de H2O desionizada. La suspensión se transfirió luego a un crisol de porcelana previamente pesado y se secó durante la noche a 105ºC y se dejó enfriar en un desecador, antes de medir el cambio de masa.
Las concentraciones de azúcares fermentables (glucosa, fructosa, maltosa y maltotriosis) se midieron por HPLC utilizando un módulo de separación de aguas 2695 y un cromatógrafo de líquidos del módulo de interfase del sistema de agua acoplado con un refractómetro diferencial de aguas 2414 (Waters Co., Milford, MA, EE. UU.). Una columna de análisis de ácido orgánico Aminex HPX-87H (300 × 7.8 mm, Bio-Rad) se equilibró con H2SO4 5 mM (Titrisol, Merck, Alemania) en agua a 55 ° C y las muestras se eluyeron con H2SO4 5 mM en agua a una temperatura Caudal de 0,3 ml / min.
Los compuestos de sabor derivados de la levadura se determinaron mediante cromatografía de gases en el espacio de cabeza con análisis de detector de ionización de llama (HS-GC-FID). Se filtraron 4 ml de muestras (0,45 µm), se incubaron a 60 ° C durante 30 minutos y luego se inyectó 1 ml de fase gaseosa (modo dividido; 225 ° C; flujo dividido de 30 ml min-1) en un cromatógrafo de gases equipado con un detector de FID y un muestreador automático de espacio de cabeza (serie Agilent 7890; Palo Alto, CA, EE. UU.). Los analitos se separaron en una columna capilar HP-5 (50 mx 320 µm x 1,05 µm columna, Agilent, EE. UU.). El gas portador fue helio (flujo constante de 1.4 mL min − 1). El programa de temperatura usó 50 ° C durante 3 min, 10 ° C min − 1 a 100 ° C, 5 ° C min – 1 a 140 ° C, 15 ° C min − 1 a 260 ° C y luego isotérmico durante 1 min. Los compuestos se identificaron por comparación con estándares auténticos y se cuantificaron utilizando el estándar.
Resultados y discusión
Generación de híbridos interespecíficos.
Se generaron con éxito mutantes auxotróficos de las cepas parentales y las cepas mutantes, A81062 ura y C12902 lys, produjeron ascosporas en el medio de esporulación sólido. Sin embargo, se observaron eficiencias de esporulación bastante bajas para la cepa ura-A81062 en comparación con la cepa lys-C12902 (11 y 78%, respectivamente). Estas ascosporas sí contenían esporas viables (12 y 71%, respectivamente) y se usaron para generar los híbridos interespecíficos. Después de aparearse y extenderse en placas de selección, las primeras colonias comenzaron a emerger después de 3 días. Después de una semana, se obtuvieron un total de 38 colonias (correspondientes a una frecuencia de hibridación de 2.6 × 10−6).
Confirmación de estado híbrido
Después de la amplificación del ADNr y la digestión con HaeIII, se obtuvo un patrón de 4 bandas para la cepa parental A81062 de S. cerevisiae, se obtuvo un patrón de 3 bandas para el S. eubayanus C12902, mientras que se obtuvieron patrones de 5 bandas para el aislado híbridos (Fig. 1a), lo que confirma que ambos genomas parentales estaban representados en estos híbridos. Este perfil RFLP de los híbridos es diferente al de los híbridos industriales de S. pastorianus que, como resultado de la pérdida de ADN posterior a la hibridación, presentan un patrón de 3 bandas o de 4 bandas dependiendo de si pertenecen al grupo de Saaz o Frohberg. [14, 31]. La amplificación de los genes FSY1 y MEX67 por PCR (Fig. 1b) también reveló la presencia de los genes de S. eubayanus y S. cerevisiae, respectivamente, en los genomas de las cepas híbridas. El estado híbrido finalmente se confirmó con PFGE, lo que también sugirió que las cepas híbridas habían heredado un conjunto completo de cromosomas de ambas cepas parentales (que se muestra en Material suplementario; Figura S1). La citometría de flujo reveló además que las cuatro cepas híbridas eran probablemente triploides (Fig. 2). Para mayor claridad, solo se muestra el perfil de fluorescencia de la cepa híbrida H1 (de las cepas híbridas) en la Fig. 2b, pero las otras cepas híbridas H2-H4 produjeron perfiles idénticos. Esto contrasta con lo que se esperaría de los apareamientos de esporas a esporas haploides, que dan como resultado la formación de células diploides. Aquí, la hibridación aparentemente fue el resultado del apareamiento raro entre una espora haploide de un padre y una célula diploide, que puede haber sufrido una pérdida de heterocigosidad y conversión a un tipo de apareamiento a / a o α / α, del otro padre [15] . Esto también explica la frecuencia de hibridación relativamente baja [1]. No se sabe qué partes de los genomas parentales se han heredado en las cepas híbridas, pero es probable que las cuatro cepas híbridas H1-H4 contengan 2n ADN de la cepa A81062 de S. cerevisiae y 1n ADN de la cepa S. eubayanus C12902 , debido a la mayor eficiencia de esporulación y la viabilidad de las esporas de C12902. Sin embargo, los datos recopilados aquí no son suficientes para mostrar esto, y también es posible que las cepas híbridas contengan proporcionalmente más ADN del padre de S. eubayanus. La proporcionalidad de la herencia de ADN podría reflejarse directamente en las propiedades funcionales de las cepas híbridas, como parece ser el caso de las levaduras Saaz y Frohberg lager [13].
 |
Figura 1
Confirmación de la hibridación mediante un rDNA ITS PCR y RFLP, y b amplificación de los genes FSY1 y MEX67. Carriles 1 y 8, escalera de ADN de 100 pb, carril 2–5 híbridos H1 – H4, carril 6 S. cerevisiae A81062 cepa parental, y carril 7 S. eubayanus C12902 cepa parental
|
 |
Figura 2
Contenido de ADN de las cepas de referencia haploides de S. cerevisiae (CEN.PK113-1A) y diploides (CEN.PK), y de la cepa híbrida H1 y cepas parentales mediante citometría de flujo. Las otras cepas híbridas H2-H4 mostraron perfiles idénticos a los del híbrido H1. Las líneas verticales discontinuas representan la intensidad de fluorescencia aproximada correspondiente al contenido de ADN 1n, 2n y 4n
|
Características de fermentación de las cepas híbridas.
Las cuatro cepas híbridas (H1-H4) fermentaron con éxito el mosto a 12 ° C y mostraron tasas de fermentación más altas que las dos cepas parentales (Fig. 3). Las fermentaciones con cepas híbridas H1 y H2 se completaron después de 8 días. Las cepas híbridas no solo fermentaron más rápido, sino que también produjeron cervezas con mayor contenido de alcohol (5,6 vs 4,5% ABV después de 11 días de fermentación). La cepa parental de S. cerevisiae A81062 creció y fermentó lentamente a 12 ° C y, por lo tanto, solo alcanzó un ABV de 4.2% después de 11 días. El crecimiento y la fermentación exitosos de las cepas híbridas a 12 ° C sugieren que la tolerancia al frío de S. eubayanus se ha transferido a las cepas híbridas. Los mecanismos que gobiernan la criotolerancia de S. eubayanus son desconocidos, pero es muy probable que esté relacionado con las diferencias en la composición de la membrana [17], así como con la actividad del producto y la expresión de los genes metabólicos centrales [29], en comparación con otras levaduras Saccharomyces. Se observaron tendencias similares en los perfiles de fermentación durante la fermentación "Generación 0" realizada a 20 ° C, ya que las cepas híbridas mostraron mejores tasas de fermentación en comparación con las dos cepas parentales (datos no mostrados). Además, las cuatro cepas híbridas y la cepa parental A81062 crecieron a 37 ° C, pero no a 40 ° C, en placas YPM, mientras que S. eubayanus C12902 no creció a ninguna temperatura, lo que sugiere que las cepas híbridas también heredaron la tolerancia al calor de la parental. La causa del fenotipo heterótico aparente de los híbridos podría ser la funcionalidad diferencial y la expresión de genes ortólogos derivados de los dos padres (específicamente ligados al metabolismo central del carbono y al transporte de azúcar), así como a diferentes valores óptimos de temperatura de los productos génicos [7, 14 ]. La pequeña diferencia en el rendimiento de la fermentación entre las cepas híbridas podría ser el resultado de la segregación meiótica durante la formación de esporas o la herencia diferencial del ADN mitocondrial, ya que el ADNmt se hereda de un solo padre [25, 35].
 |
Fig. 3
a El contenido de alcohol (% ABV), b peso específico yc la masa seca de levadura suspendida (g / L) del mosto a 12 ° P fermentado con las cepas híbridas (líneas continuas) y las cepas parentales (líneas de puntos). Los valores son medias de dos fermentaciones independientes y barras de error donde visibles representan el rango
|
Los perfiles de azúcar del mosto original y las cervezas (Tabla 1) revelan que la mayor atenuación alcanzada por las cepas híbridas en comparación con el progenitor de S. eubayanus es el resultado de la utilización diferencial de la maltotriosa. La capacidad de fermentar la maltotriosa parece haber sido heredada de la matriz de S. cerevisiae, ya que los ensayos de transporte de azúcar activo confirmaron que todas las cepas híbridas y S. cerevisiae A81062 eran capaces de transportar este azúcar a través de la membrana celular, mientras que S. eubayanus C12902 mostraron una captación insignificante (Tabla 2) como se mostró anteriormente por Gibson et al. [13]. La captación de maltotriosa es común en las cepas de levadura lager; sin embargo, esta capacidad parece estar limitada a las cepas que pertenecen al grupo Frohberg [13]. El uso de maltosa y maltotriosa durante la fermentación depende de la actividad de un rango de transportadores transmembrana [46]. En las cepas de cerveza industrial, la maltotriosa es transportada esencialmente a la célula por los transportadores AGT1, mientras que en las cepas de levadura lager los genes AGT1 derivados de S. cerevisiae permanecen no funcionales, y se cree que el transporte de la maltotriosa se realiza por Mtt1 ( Mty1) y una forma divergida de AGT1, que presumiblemente han sido heredadas del padre de S. eubayanus [10, 45, 46, 48]. Esto contradice el hecho de que aquí y en estudios previos [13] se observó un transporte insignificante de maltotriosa a la cepa de tipo S. eubayanus, pero el uso de la maltotriosa de otras cepas de S. eubayanus, p. Ej. De diferentes orígenes geográficos, en la actualidad permanece inexplorado. La utilización de la maltotriosa es importante durante las fermentaciones de la cervecería, ya que es el segundo azúcar más abundante en el mosto, su utilización da como resultado un mayor rendimiento de alcohol y la maltotriosa residual puede afectar el sabor [51]. Los perfiles de fermentación (Fig. 3a, b) revelan que las cepas híbridas y S. eubayanus mostraron una tasa de fermentación similar al comienzo de la fermentación, cuando los monosacáridos son predominantemente absorbidos y utilizados por la levadura [3]. Después de las primeras 48 h, las tasas de fermentación de las cepas híbridas aumentan en relación con S. eubayanus, probablemente debido a la utilización más eficiente de maltosa y maltotriosa en las primeras. Las cepas híbridas y S. eubayanus mostraron tasas de fermentación mayores que las de S. cerevisiae durante prácticamente toda la fermentación porque la temperatura es claramente subóptima para S. cerevisiae. Esto también es evidente por el hecho de que había maltosa residual en la cerveza fermentada con S. cerevisiae (Tabla 1). Aquí, se usó un mosto de gravedad relativamente baja (12 ° Plato), pero la industria está mostrando interés hacia obras de gravedad más alta [37]. Si bien el rendimiento de la fermentación de las cepas híbridas en el mosto de alta gravedad no se probó en este estudio, es posible que posean una mayor tolerancia al estrés como resultado de su poliploidía [52].
Tabla 1
Azúcares (g / L) en el mosto original y cervezas fermentadas con las cepas híbridas y parentales
Yeast strain/sample
|
Maltose
|
Maltotriose
|
|---|
Original wort
|
59.1 (±0.37)
|
18.8 (±0.21)
|
Hybrid H1
|
1.6 (±0.04)
|
3.6 (±0.07)
|
Hybrid H2
|
1.5 (±0.03)
|
3.4 (±0.05)
|
Hybrid H3
|
1.6 (±0.02)
|
3.9 (±0.13)
|
Hybrid H4
|
1.6 (±0.20)
|
5.3 (±0.24)
|
A81062
|
25.1 (±0.69)
|
5.5 (±0.09)
|
C12902
|
1.8 (±0.15)
|
18.5 (±0.58)
|
Yeast strain
|
Flocculation ability (%)
|
Maltose uptake µmol min−1 g−1 DY (5 mM maltose)
|
Maltotriose uptake µmol min−1 g−1 DY (5 mM maltotriose)
|
|---|
Hybrid H1
|
85 (±2.1)
|
12.6 (±1.7)
|
4.1 (±0.3)
|
Hybrid H2
|
87 (±1.3)
|
12.3 (±3.0)
|
4.1 (±0.2)
|
Hybrid H3
|
88 (±1.6)
|
15.8 (±0.9)
|
5.4 (±1.4)
|
Hybrid H4
|
82 (±4.5)
|
16.1 (±0.7)
|
5.4 (±1.3)
|
A81062
|
71 (±4.2)
|
10.1 (±0.5)
|
4.2 (±0.2)
|
C12902
|
15 (±0.8)
|
13.2 (±3.1)
|
0.1 (±0.0)
|
Las concentraciones de compuestos aromáticos en las cervezas (Fig. 4) revelan que las cuatro cepas híbridas produjeron cervezas con perfiles de aroma similares. En comparación con las cepas híbridas, S. eubayanus C12902 produjo más alcoholes superiores relacionados con la síntesis de aminoácidos de cadena ramificada (casi dos concentraciones de 2-metilpropanol, 3-metilbutanol y 2-metilbutanol). Éstos le dan a la cerveza aromas alcohólicos y similares a solventes, que generalmente se consideran desagradables [32]. A pesar de estas dos diferencias en las concentraciones de 3-metilbutanol, las concentraciones de acetato de 3-metilbutilo fueron bastante similares para las cepas híbridas y S. eubayanus (alrededor del umbral de sabor de 1.2 mg / L [26]). El acetato de 3-metilbutilo le da a la cerveza un aroma parecido al plátano y la pera, que se considera deseable en varios estilos de cerveza [32]. Las cepas híbridas produjeron más ésteres etílicos que ambas cepas parentales, y las concentraciones de hexanoato de etilo superaron el umbral de sabor de 0.2 mg / L [26]. Los ésteres etílicos le dan a la cerveza un aroma afrutado y parecido a una manzana. La causa de estas diferencias en la formación de ésteres de las cepas híbridas en comparación con ambas cepas parentales no se conoce, pero podría deberse a diferencias genéticas (por ejemplo, aumento de la expresión y funcionalidad diferente de los genes ortólogos [5, 37, 44]) y un efecto indirecto. Resultado de las fermentaciones, p. ej. por la formación de más alcohol y los precursores de ácidos grasos, las diferencias en el pH del mosto o las diferencias en el crecimiento de la levadura [18, 49]. La formación de ésteres de acetato depende principalmente de la expresión y de las actividades enzimáticas de los genes ATF1 y ATF2 que codifican transferasa [44], mientras que Saerens et al. [36, 37] encontraron que las enzimas codificadas por los genes EHT1 y EEB1 son las principales responsables de la síntesis del éster etílico. En general, se observó una baja cantidad de ésteres en la cerveza fermentada con S. cerevisiae A81062. Este fue el resultado más probable de la temperatura de fermentación baja y subóptima, que resultó en una menor expresión y actividad enzimática de los genes involucrados en la síntesis de ésteres, p. ATF1, ATF2, EEB1 y EHT1. Se ha demostrado que la expresión de estos genes, que codifican las enzimas transferasa involucradas en la síntesis de acetato y éster etílico, aumenta a temperaturas crecientes [37, 44]. Los resultados sugieren que podría ser posible aumentar la diversidad aromática de las cepas de levadura lager a través de la hibridación interespecífica y producir cepas de levadura lager con la producción de aroma deseada a través de la selección de cepas parentales apropiadas. Sin embargo, aún se debe hacer más trabajo sobre el tema, especialmente con respecto a las relaciones entre la expresión génica ortóloga y la producción de aroma. Todas las cervezas también tenían un aroma parecido al de un clavo de olor, causado por la presencia de 4-vinilguaiacol [8], lo que sugiere que las cepas híbridas habían heredado el gen PAD1, que codifica la enzima descarboxilasa del ácido fenilacrílico responsable de la conversión del ácido ferúlico a 4 -vinilguaiacol, de cualquiera de los padres, que eran ambos PAD1 + (nuestros propios datos no publicados).
 |
Fig. 4
Las concentraciones de compuestos aromáticos en las cervezas fermentadas con las cepas híbridas y parentales (mg / L). Donde está visible, la línea discontinua representa el umbral de sabor típico [26]. Los valores son medias de dos fermentaciones independientes y barras de error donde visibles representan el rango
|
El ensayo de floculación reveló una fuerte floculación tanto para la cepa parental de S. cerevisiae A81062 como para las cuatro cepas híbridas H1-H4, mientras que S. eubayanus C12902 mostró una floculación deficiente (Tabla 2). Estas propiedades también fueron evidentes durante la fermentación, ya que se observó más biomasa suspendida en las fermentaciones con S. eubayanus que con los híbridos (Fig. 3c). La capacidad de floculación de la levadura se define principalmente por la forma en que los genes FLO son funcionales en varias cepas [47], y en este caso las cepas híbridas tienen una capacidad de floculación heredada claramente del progenitor de S. cerevisiae. Sorprendentemente, la floculación de las cepas híbridas fue más fuerte que las cepas parentales, revelando así una amplificación del fenotipo (Tabla 2). La floculación de la levadura hacia el final de la fermentación permite una forma barata y efectiva de eliminar la levadura de la cerveza, y el uso de cepas de levadura fuertemente floculantes es especialmente popular entre las cervecerías más pequeñas.
En conclusión, el uso del apareamiento en masa para generar híbridos de levadura lager novo a través de la hibridación interespecífica entre una cepa de S. cerevisiae ale y S. eubayanus fue exitoso. Las cepas híbridas resultantes no solo heredaron propiedades beneficiosas de ambas cepas parentales (criotolerancia, utilización de maltotriosa y fuerte floculación), sino que también mostraron un vigor híbrido aparente en comparación con las cepas parentales al fermentar más rápido y lograr una utilización más completa de los azúcares fermentables. Debe mencionarse que aquí, el rendimiento de las cepas híbridas se comparó en relación con las cepas parentales originales y no con el producto de esporas haploides reales que se aparearon para formar los híbridos triploides. La posible heterocigosidad de las cepas parentales sugiere que los productos de esporas pueden ser genéticamente muy diferentes de cualquier otro producto de esporas y, por lo tanto, también tienen un comportamiento de fermentación bastante diferente al de las esporas parentales u otras esporas hermanas [2, 41]. Todavía no se sabe cómo se crearon por primera vez los híbridos de levadura lager natural. El rendimiento claramente mejorado de la fermentación de las cepas híbridas mostradas aquí apoyaría la hipótesis de que S. eubayanus pudo haber estado inicialmente presente como un contaminante en las fermentaciones de ale de S. cerevisiae. A temperaturas más bajas, el estado híbrido, que confiere beneficios fenotípicos de ambos progenitores, se habría seleccionado y los híbridos habrían dominado rápidamente el proceso de elaboración [14, 42]. También se sabe que los genomas de los híbridos de levadura recién creados son inestables, y esto puede permitir la adaptación evolutiva de los híbridos a diferentes ambientes [20]. Estos resultados sugieren que la hibridación interespecífica es adecuada para la producción de nuevas cepas de levadura lager no GM con propiedades únicas (por ejemplo, la producción de sabor o la tolerancia elevada al estrés) y la generación de nuevos estilos de cerveza. Se requiere una investigación adicional de las propiedades de los híbridos de novo, especialmente en relación con la proporcionalidad de la herencia del ADN, el origen del ADN mitocondrial y la producción de aromas, y puede ayudar a dilucidar la historia evolutiva de las cepas de levadura industriales. Como observación final, queremos mencionar el trabajo reciente de Marit Hebly y colaboradores de Delft University of Technology titulado "Saccharomyces cerevisiae x Saccharomyces eubayanus interespecífico híbrido, lo mejor de ambos mundos y más allá" que refuerza nuestras observaciones y complementa idealmente nuestro estudio. Este trabajo será publicado en breve en FEMS Yeast Research.
Notas
Expresiones de gratitud
Agradecemos a Annika Wilhelmson por su apoyo en todo el proceso, a Eero Mattila y Arvi Wilpola por la preparación del mosto y otra asistencia en la fábrica de cerveza VTT Pilot, ya Aila Siltala y Sirpa Jylhä por su asistencia técnica especializada. Este trabajo fue apoyado por la Fundación Alfred Kordelin, el Laboratorio de elaboración de cerveza PBL, la Academia de Finlandia (Proyecto de la Academia 276480) y el 7PM Marie-Curie ITN YEASTCELL.
Material suplementario
10295_2015_1597_MOESM1_ESM.pdf (1.1 mb)
Material complementario 1 (PDF 1123 kb)
Referencias
1.
Bellon J, Eglinton J, Siebert T, Pollnitz A, Rose L, de Barros Lopes M, Chambers P (2011) Newly generated interspecific wine yeast hybrids introduce flavour and aroma diversity to wines. Appl Microbiol Biotechnol 91:603–612
CrossRefPubMedGoogle Scholar
2.
Bilinski C, Russell I, Stewart G (1986) Analysis of sporulation in brewer’s yeast: induction of tetrad formation. J Inst Brew 92:594–598
CrossRefGoogle Scholar
3.
Bisson L, Coons D, Kruckeberg A, Lewis D (1993) Yeast sugar transporters. Crit Rev Biochem Mol Biol 28:259–308
CrossRefPubMedGoogle Scholar
4.
Boeke J, Trueheart J, Natsoulis G, Fink G (1987) 5-Fluoroorotic acid as a selective agent in yeast molecular genetics. Methods Enzymol 154:164–175
CrossRefPubMedGoogle Scholar
5.
Bolat I, Romagnoli G, Zhu F, Pronk J, Daran J (2013) Functional analysis and transcriptional regulation of two orthologs of ARO10, encoding broad-substrate-specificity 2-oxo-acid decarboxylases, in the brewing yeast
Saccharomyces pastorianusCBS1483. FEMS Yeast Res 13:505–517
CrossRefPubMedGoogle Scholar
6.
Cebollero E, Gonzalez-Ramos D, Tabera L, Gonzalez R (2007) Transgenic wine yeast technology comes of age: is it time for transgenic wine? Biotechnol Lett 29:191–200
CrossRefPubMedGoogle Scholar
8.
Coghe S, Benoot K, Delvaux F, Vanderhaegen B, Delvaux FR (2004) Ferulic acid release and 4-vinylguaiacol formation during brewing and fermentation: indication for feruloyl esterase activity in
Saccharomyces cerevisiae. J Agric Food Chem 52:602–608
CrossRefPubMedGoogle Scholar
9.
D’Hautcourt O, Smart K (1999) Measurement of brewing yeast flocculation. J Am Soc Brew Chem 57:123–128
Google Scholar
10.
Dietvorst J, Londesborough J, Steensma HY (2005) Maltotriose utilization by lager yeast strains: MTT1 encodes a maltotriose transporter. Yeast 22:775–788
CrossRefPubMedGoogle Scholar
12.
Garcia Sanchez R, Solodovnikova N, Wendland J (2012) Breeding of lager yeast with
Saccharomyces cerevisiae improves stress resistance and fermentation performance. Yeast 29:343–355
CrossRefPubMedGoogle Scholar
13.
Gibson B, Storgårds E, Krogerus K, Vidgren V (2013) Comparative physiology and fermentation performance of Saaz and Frohberg lager yeast strains and the parental species
Saccharomyces eubayanus. Yeast 30:255–266
CrossRefPubMedGoogle Scholar
14.
Gibson B, Liti G (2015)
Saccharomyces pastorianus: genomic insights inspiring innovation for industry. Yeast 32:17–27
PubMedGoogle Scholar
15.
Gunge N, Nakatomi Y (1972) Genetic mechanisms of rare matings of the yeast
Saccharomyces cerevisiae heterozygous for mating type. Genetics 70:41–58
PubMedCentralPubMedGoogle Scholar
16.
Haase S, Reed S (2002) Improved flow cytometric analysis of the budding yeast cell cycle. Cell Cycle 1:132–136
CrossRefPubMedGoogle Scholar
17.
Henderson C, Lozada-Contreras M, Jiranek V, Longo M, Block D (2013) Ethanol production and maximum cell growth are highly correlated with membrane lipid composition during fermentation as determined by lipidomics analysis of 22
Saccharomyces cerevisiae strains. Appl Environ Microbiol 79:91–104
CrossRefPubMedCentralPubMedGoogle Scholar
18.
Hiralal L, Olaniran A, Pillay B (2014) Aroma-active ester profile of ale beer produced under different fermentation and nutritional conditions. J Biosci Bioeng 117:57–64
CrossRefPubMedGoogle Scholar
19.
Krogerus K, Gibson B (2013) Influence of valine and other amino acids on total diacetyl and 2,3-pentanedione levels during fermentation of brewer’s wort. Appl Microbiol Biotechnol 97:6919–6930
CrossRefPubMedCentralPubMedGoogle Scholar
20.
Kunicka-Styczynska A, Rajkowska K (2011) Physiological and genetic stability of hybrids of industrial wine yeasts
Saccharomyces sensu stricto complex. J Appl Microbiol 110:1538–1549
CrossRefPubMedGoogle Scholar
21.
Libkind D, Hittinger C, Valerio E, Goncalves C, Dover J, Johnston M, Goncalves P, Sampaio J (2011) Microbe domestication and the identification of the wild genetic stock of lager-brewing yeast. Proc Natl Acad Sci USA 108:14539–14544
CrossRefPubMedCentralPubMedGoogle Scholar
23.
Liti G, Peruffo A, James S et al (2005) Inferences of evolutionary relationships from a population survey of LTR-retrotransposons and telomeric-associated sequences in the
Saccharomyces sensu stricto complex. Yeast 22:177–192
CrossRefPubMedGoogle Scholar
24.
Lucero P, Herweijer M, Lagunas R (1993) Catabolite inactivation of the yeast maltose transporter is due to proteolysis. FEBS Lett 333:165–168
CrossRefPubMedGoogle Scholar
25.
Marullo P, Bely M, Masneuf-Pomarede I, Aigle M, Dubourdieu D (2004) Inheritable nature of enological quantitative traits is demonstrated by meiotic segregation of industrial wine yeast strains. FEMS Yeast Res 4:711–719
CrossRefPubMedGoogle Scholar
26.
Meilgaard M (1982) Prediction of flavor differences between beers from their chemical composition. J Agric Food Chem 30:1009–1017
CrossRefGoogle Scholar
27.
Merritt A, Hurley R (1972) Evaluation of sporulation media for yeasts obtained from pathological material. J Med Microbiol 5:21–30
CrossRefPubMedGoogle Scholar
28.
Muir A, Harrison E, Wheals A (2011) A multiplex set of species-specific primers for rapid identification of members of the genus
Saccharomyces. FEMS Yeast Res 11:552–563
CrossRefPubMedGoogle Scholar
29.
Paget C, Schwartz J, Delneri D (2014) Environmental systems biology of cold-tolerant phenotype in
Saccharomyces species adapted to grow at different temperatures. Mol Ecol 23:5241–5257
CrossRefPubMedCentralPubMedGoogle Scholar
30.
Pengelly R, Wheals A (2013) Rapid identification of
Saccharomyces eubayanus and its hybrids. FEMS Yeast Res 13:156–161
CrossRefPubMedGoogle Scholar
31.
Pham T, Wimalasena T, Box W, Koivuranta K, Storgårds E, Smart K, Gibson B (2011) Evaluation of ITS PCR and RFLP for differentiation and identification of brewing yeast and brewery ‘wild’ yeast contaminants. J Inst Brew 117:556–568
CrossRefGoogle Scholar
32.
Pires E, Teixeira J, Branyik T, Vicente A (2014) Yeast: the soul of beer’s aroma—a review of flavour-active esters and higher alcohols produced by the brewing yeast. Appl Microbiol Biotechnol 98:1937–1949
CrossRefPubMedGoogle Scholar
34.
Powell C, Casalis F, Fischborn T (2012) Stress tolerance in group 1 and group 2 lager brewing yeasts. In: Proceedings of the World Brewing Congress 2012, Portland, OR, USA July 28–August 1, 2012
Google Scholar
35.
Rainieri S, Kodama Y, Nakao Y, Pulvirenti A, Giudici P (2008) The inheritance of mtDNA in lager brewing strains. FEMS Yeast Res 8:586–596
CrossRefPubMedGoogle Scholar
36.
Saerens D, Verstrepen K, Van Laere S, Voet A, Van Dijck P, Delvaux FR, Thevelein J (2006) The
Saccharomyces cerevisiae EHT1 and
EEB1 genes encode novel enzymes with medium-chain fatty acid ethyl ester synthesis and hydrolysis capacity. J Biol Chem 281:4446–4456
CrossRefPubMedGoogle Scholar
37.
Saerens S, Verbelen P, Vanbeneden N, Thevelein J, Delvaux FR (2008) Monitoring the influence of high-gravity brewing and fermentation temperature on flavour formation by analysis of gene expression levels in brewing yeast. Appl Microbiol Biotechnol 80:1039–1051
CrossRefPubMedGoogle Scholar
38.
Sato M, Kishimoto M, Watari J, Takashio M (2002) Breeding of brewer’s yeast by hybridization between a top-fermenting yeast
Saccharomyces cerevisiae and a cryophilic yeast
Saccharomyces bayanus. J Biosci Bioeng 93:509–511
CrossRefPubMedGoogle Scholar
39.
Steensels J, Meersman E, Snoek T, Saels V, Verstrepen K (2014) Large-scale selection and breeding to generate industrial yeasts with superior aroma production. Appl Environ Microbiol. doi:
10.1128/AEM.02235-14PubMedCentralGoogle Scholar
40.
Stewart G, Hill A, Russell I (2013) 125th anniversary review: developments in brewing and distilling yeast strains. J Inst Brew 119:202–220
CrossRefGoogle Scholar
41.
Tsuboi M, Takahashi T (1988) Genetic analysis of the non-sporulating phenotype of brewer’s yeasts. J Ferment Technol 66:605–613
CrossRefGoogle Scholar
43.
van Dijken J, Bauer J, Brambilla L et al (2000) An interlaboratory comparison of physiological and genetic properties of four
Saccharomyces cerevisiae strains. Enzyme Microbiol Technol 26:706–714
CrossRefGoogle Scholar
44.
Verstrepen K, Van Laere S, Vanderhaegen B, Derdelinckx G, Dufour J, Pretorius I, Winderickx J, Thevelein J, Delvaux FR (2003) Expression levels of the yeast alcohol acetyltransferase genes ATF1, Lg-ATF1, and
ATF2 control the formation of a broad range of volatile esters. Appl Environ Microbiol 69:5228–5237
CrossRefPubMedCentralPubMedGoogle Scholar
45.
Vidgren V, Ruohonen L, Londesborough J (2005) Characterization and functional analysis of the MAL and MPH loce for maltose utilization in some ale and lager yeast strains. Appl Environ Microbiol 71:7846–7857
CrossRefPubMedCentralPubMedGoogle Scholar
46.
Vidgren V, Huuskonen A, Virtanen H, Ruohonen L, Londesborough J (2009) Improved fermentation performance of a lager yeast after repair of its
AGT1 maltose and maltotriose transporter genes. Appl Environ Microbiol 75:2333–2345
CrossRefPubMedCentralPubMedGoogle Scholar
47.
Vidgren V, Londesborough J (2011) 125th anniversary review: yeast flocculation and sedimentation in brewing. J Inst Brew 117:475–487
CrossRefGoogle Scholar
48.
Vidgren V, Londesborough J (2012) Characterization of the
Saccharomyces bayanus-type
AGT1 transporter of lager yeast. J Inst Brew 118:148–151
CrossRefGoogle Scholar
49.
Yoshioka K, Hashimoto N (1981) Ester formation by alcohol acetyltransferase from brewer’s yeast. Agric Biol Chem 45:2183–2190
CrossRefGoogle Scholar
50.
Zaret K, Sherman F (1985) Alpha-aminoadipate as a primary nitrogen source for
Saccharomyces cerevisiae mutants. J Bacteriol 162:579–583
PubMedCentralPubMedGoogle Scholar
51.
Zheng X, D’Amore T, Russell I, Stewart G (1994) Factors influencing maltotriose utilization during brewery wort fermentations. J Am Soc Brew Chem 52:41–47
Google Scholar
Material suplementario
Figura S1. Separación PFGE de cromosomas de cepas híbridas H1-H4 y cepas parentales. Carril 1 y 8 marcador cromosómico cepa YNN295, carril 2-5 híbridos H1-H4, carril 6 S. cerevisiae A81062 cepa parental, y carril 7 S. eubayanus C12902 cepa parental. Los cromosomas se identifican a la izquierda: los cromosomas VII y XV no se resuelven; el cromosoma II viaja inmediatamente arriba
cromosoma xiv.
© The Author(s) 2015
Open AccessThis article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
- Cite this article as:
- Krogerus, K., Magalhães, F., Vidgren, V. et al. J Ind Microbiol Biotechnol (2015) 42: 769. https://doi.org/10.1007/s10295-015-1597-6
Published in cooperation with
Society for Industrial Microbiology and Biotechnology