Volumen 24, Número 10, 19 de mayo de 2014 , páginas R380-R381
JianBing1 2 Pei-JieHan 1 Wan-QiuLiu 1 2 Qi-MingWang 1 Feng-YanBai 1

La elaboración de cerveza lager surgió en el siglo XV en Baviera [1] y actualmente es la técnica más popular para la producción de bebidas alcohólicas en el mundo. La técnica se caracteriza por una fermentación a baja temperatura utilizando la levadura doméstica Saccharomyces pastorianus (sinónimo S. carlsbergensis ). Ha quedado claro que la levadura lager es un híbrido con una parte de su genoma que se originó a partir de la levadura ale de S. cerevisiae [2] . Sin embargo, la fuente del subgenoma no-ale, que otorga a la levadura lager una tolerancia al frío, ha sido un tema de debate [3] . Recientemente, se ha propuesto una hipótesis de origen patagónico de levadura lager basada en el descubrimiento de un nuevo Saccharomyces criotolerante.Especies de los bosques nativos patagónicos de argentina [4] . Esta levadura, llamada S. eubayanus, exhibió la concordancia más cercana conocida (99.56%) a la porción de no-ale de la levadura lager y, por lo tanto, se creía que era su progenitora. Sin embargo, ahora mostramos que esta especie de levadura es probablemente nativa de la meseta tibetana. Una de las poblaciones tibetanas de la especie muestra una afinidad más cercana con la levadura lager que la población patagónica, como se deduce de la genética de la población y los análisis de la secuencia del genoma. Por lo tanto, proporcionamos pruebas sólidas de una hipótesis de origen del Lejano Oriente de la levadura lager, que aparentemente se corresponde mejor con la geografía y la historia del comercio mundial.
Texto principal
S. eubayanus no se ha encontrado en otras regiones del mundo, a pesar de más de un siglo de investigación sobre levaduras en ciernes en todo el mundo y un estudio extenso reciente con un esfuerzo especial para aislar especies de Saccharomyces criotolerantes de ambientes arbóreos en Europa [5] . Como la reserva genética silvestre de levadura lager, la ausencia de S. eubayanus en Europa es sorprendente, lo que lleva a la propuesta de que S. eubayanus puede haber sido importado de la Patagonia después de la llegada del comercio transatlántico [4]. Aunque esta hipótesis ha generado un gran interés entre los científicos, los cerveceros y el público, varios puntos del escenario siguen siendo desconcertantes. Lo más importante es que las líneas de tiempo de los eventos históricos relacionados entran en conflicto con esta hipótesis. Lager-brewing fue inventado en el 1400 en Baviera [1] , mientras que el comercio transatlántico comenzó más tarde en el 1500 después del primer viaje de Colón al nuevo mundo.
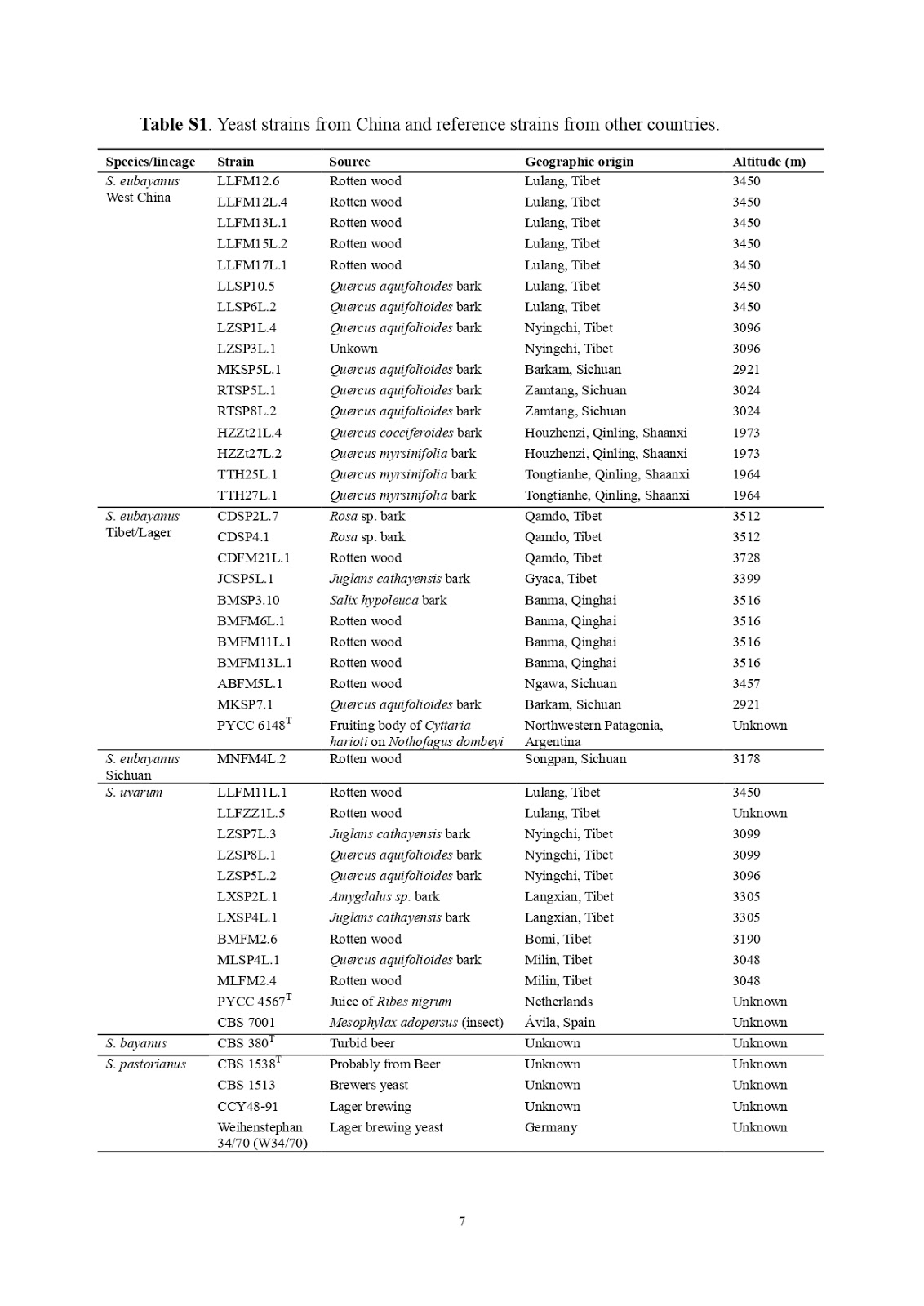
En los últimos años, hemos aislado con éxito S. eubayanus y su pariente cercano S. uvarum de la corteza y madera podrida de diferentes robles y otros árboles de hoja caduca recolectados de la meseta tibetana, incluidas áreas de gran altitud en el Tíbet, Qinghai y Sichuan en el oeste China y de la montaña Qingling en la provincia de Shaanxi, una región límite entre subtropical y templado cálido en el noroeste de China (información complementaria). El análisis filogenético molecular basado en múltiples loci mostró que, además de S. uvarum , existen en China al menos tres linajes distintos de S. eubayanus : primero, el linaje Tíbet / Lager que incluye 10 cepas de la meseta tibetana, el tipo de cepa (PYCC) 6148 T) de las especies de las cepas Patagonia y S. pastorianus (CCY48-91 y Weihenstephan 34/70); segundo, el linaje del oeste de China consistía en 16 cepas de diferentes regiones del oeste de China; y tercero, el linaje de Sichuan representado por una sola cepa de un área marginal de la meseta tibetana en la provincia de Sichuan ( Figura 1 ; Información complementaria). Se observó una clara congruencia genealógica para los tres linajes de S. eubayanus y el linaje de S. uvarum , pero no para la cepa de tipo híbrido de S. bayanus (información complementaria). Cruces interlineales de S. eubayanusmostraron 18 a 37% (31,1% en promedio) de viabilidad de esporas (información complementaria), lo que sugiere que estos linajes representan diferentes poblaciones de una especie con aislamiento reproductivo parcial. La viabilidad de las esporas de los cruzamientos entre poblaciones y entre especies no fue clara (información complementaria), lo que desafió la separación de S. eubayanus y S. uvarum a nivel de especie [4] . El descubrimiento de S. eubayanus y S. uvarum en este estudio y otras especies de Saccharomyces en estudios anteriores [6] sugiere que el Lejano Oriente asiático parece ser la única área en el mundo que alberga todas las especies biológicas reconocidas de Saccharomyces [7], apoyando la hipótesis de que el Lejano Oriente asiático es el posible centro de origen de las levaduras Saccharomyces 6 , 8 .
 |
| Figura 1. Filogenia de las levaduras lager.
Árbol filogenético construido a partir del análisis bayesiano de las secuencias concatenadas de nueve genes de proteínas (CCA1, FUN14, FSY1, GDH1, HIS3, MET2, MLS1, PDR10 y RIP1) y tres regiones intergénicas (entre APP1 e YPT53; FAR8 y RSF1; y MSL1 y DSN1, respectivamente) con una longitud total de 10.657 pb, que muestran las relaciones entre los linajes de S. eubayanus, S. uvarum y S. bayanus. La raíz se establece en el punto medio. Las longitudes de las ramas se escalan en términos de números esperados de sustituciones de nucleótidos por sitio. Las probabilidades posteriores del análisis bayesiano / porcentajes de arranque de 1000 réplicas del análisis de máxima verosimilitud se muestran en las ramas principales.
|
Las cepas chinas de S. eubayanus mostraron una diversidad de secuencia (ϕ) de 3.938 × 10-2, calculada a partir de los SNP reconocidos en los 12 loci secuenciados. Los tres linajes de S. eubayanus divergieron entre sí y del linaje de S. uvarum en 6.02–7.57% y 9.28–10.31% de variaciones de secuencia, respectivamente. La diversidad genética de S. eubayanus en China parece extremadamente alta, en comparación con otra especie silvestre de S. paradoxus, que es cosmopolita, con una población bien estructurada delineada a lo largo de los límites geográficos [9] y con un grado de aislamiento productivo inter-linaje similar al de S. eubayanus [10]. Las poblaciones globales de S. paradoxus mostraron una diversidad general de solo 1.638 × 10-2 y divergencias interpoblacionales de solo 1.4–4.2%, calculadas a partir de un conjunto similar de 13 loci [6]. Estos datos sugieren que S. eubayanus es muy probablemente una especie nativa de China.
El árbol filogenético también mostró que la población tibetana del linaje S. eubayanus Tibet / Lager estaba más estrechamente relacionada con S. pastorianus que con el tipo de cepa PYCC 6148T de Patagonia (Figura 1). En los 12 loci comparados, las identidades de secuencia global de las cepas tibetanas y la cepa patagónica con S. pastorianus Weihenstephan 34/70 fueron 99.77–99.82% y 99.35%, respectivamente. La afinidad más cercana de las cepas tibetanas con S. pastorianus también se mostró en todos los árboles genéticos individuales (Figura S3). La identidad media de la secuencia del genoma completo de una cepa tibetana con el resto no-ale de S. pastorianus Weihenstephan 34/70 fue del 99,82% (información complementaria), siendo más alta que la identidad de la secuencia del genoma del 99,56% entre la cepa patagónica PYCC 6148T y Weihenstephan 34 / 70, como se informó en Libkind et al. [4] basado en lecturas de Illumina mucho más cortas (36 pb) con una profundidad de secuenciación del genoma de solo 13 veces la cobertura.
Nuestros resultados sugieren fuertemente que la población tibetana de S. eubayanus es el donante directo de los no-S. Cerevisiae subgenoma de la levadura lager. Europa y Asia están ubicadas en la misma masa de tierra, y habría sido mucho más fácil para las cepas tibetanas de S. eubayanus llegar a Europa a través del puente continental euroasiático. La historia del comercio entre Asia y Europa a través de la Ruta de la Seda se inició hace aproximadamente 2000 años. Hubiera habido mucho tiempo para que las cepas tibetanas de S. eubayanus colonizaran Europa antes de ser domesticadas para la elaboración de cerveza en Baviera en el siglo XV. Sin embargo, cuándo y cómo las cepas de S. eubayanus del Lejano Oriente se abrieron camino a Sudamérica quedan por dilucidar.
Expresiones de gratitud
Agradecemos a José Paulo Sampaio y Teun Boekhout por proporcionar cepas de referencia; y Richard C. Gardner, Chris T. Hittinger y Gennadi I. Naumov por sus comentarios críticos sobre las primeras versiones del manuscrito. Este estudio fue apoyado por la subvención no. 91131004 de la Fundación Nacional de Ciencias Naturales de China y no. 2012FY111600 del Ministerio de Ciencia y Tecnología de China.
Información suplementaria
Descargar el archivo PDF Acrobat (253KB)
Documento S1. Procedimientos experimentales tres tablas y tres figuras.
Referencias
1 H.S. CorranA History of Brewing
David and Charles, London (1975)
2 Y. Nakao, T. Kanamori, T. Itoh, Y. Kodama, S. Rainieri, N. Nakamura, T. Shimonaga, M. Hattori, T. AshikariGenome sequence of the lager brewing yeast, an interspecies hybrid
DNA Res., 16 (2009), pp. 115-129
3 B. Dunn, G. SherlockReconstruction of the genome origins and evolution of the hybrid lager yeast Saccharomyces pastorianus
Genome Res., 18 (2008), pp. 1610-1623
4 D. Libkind, C.T. Hittinger, E. Valério, C. Gonçalves, J.Dover, M. Johnston, P. Gonçalves, J.P. SampaioMicrobe domestication and the identification of the wild genetic stock of lager-brewing yeast
Proc. Natl. Acad. Sci. USA, 108 (2011), pp. 14539-14544
5 J.P. Sampaio, P. GonçalvesNatural populations of Saccharomyces kudriavzevii in Portugal are associated with oak bark and are sympatric with S. cerevisiae and S. paradoxus
Appl. Environ. Microbiol., 74 (2008), pp. 2144-2152
6 Q.M. Wang, W.Q. Liu, G. Liti, S.A. Wang, F.Y. BaiSurprisingly diverged populations of Saccharomyces cerevisiae in natural environments remote from human activity
Mol. Ecol., 21 (2012), pp. 5404-5417
7 C.T. HittingerSaccharomyces diversity and evolution: a budding model genus
Trends Genet., 29 (2013), pp. 309-317
8 G.I. Naumov, T.A. NikonenkoThe East Asia is a probable land of the cultured yeasts Saccharomyces cerevisiae (in Russian)
Izv. Sib. Otd. Akad. Nauk SSSR Ser. Biol. Nauk, 20 (1988), pp. 97-101
9 G. Liti, D.M. Carter, A.M. Moses, J. Warringer, L. Parts, S.A. James, R.P. Davey, I.N. Roberts, A. Burt, V.Koufopanou, et al.Population genomics of domestic and wild yeasts
Nature, 458 (2009), pp. 337-341
10 H.A. Kuehne, H.A. Murphy, C.A. Francis, P.D.SniegowskiAllopatric divergence, secondary contact, and genetic isolation in wild yeast populations
Curr. Biol., 17 (2007), pp. 407-411